Rédacteur : Jean Terrisse
Physionomie – écologie
Cet habitat générique englobe divers types de boisements caducifoliés non hygrophiles, généralement dominés par une essence unique (rarement plus), appartenant au fonds dendrologique indigène ou naturalisée depuis longtemps (Castanea, Robinia) et formant généralement un sylvofaciès clairement distinct au sein des massifs forestiers régionaux. Les essences plantées et/ou nécessitant une intervention humaine plus ou moins soutenue pour leur survie (peupliers par ex.) ne sont pas incluses ici et font l’objet d’une fiche spécifique (PLANTATIONS DE FEUILLUS).
La physionomie, la structure et l’écologie de ces peuplements étant très variables, celles-ci seront examinées successivement en fonction de chacun des 6 principaux types distingués dans la région :
– les bois de Châtaignier : le Châtaignier Castanea sativa n’est pas une essence indigène dans l’ouest atlantique (arbre originaire des basses montagnes du pourtour méditerranéen) mais il y a été introduit dans les régions à hiver doux depuis très longtemps (époque romaine) pour la valeur calorique de ses fruits (châtaignes) et les multiples usages de son bois. Si l’usage alimentaire est devenu marginal de nos jours, la croissance rapide de l’espèce et sa capacité à rejeter de souche en font encore une essence largement plantée et exploitée, souvent en substitution de forêts acidiphiles (20 000ha en Poitou-Charentes en 2009). Il prospère sur des sols pauvres en calcaire et en bases, souvent sableux ou sablo-limoneux, ni trop frais ni trop secs. Les châtaigneraies régionales ont en général l’aspect d’un taillis peu élevé (10-15m), homogène quant aux classes d’âge, d’où émergent parfois 1 ou 2 grands Quercus témoins de la forêt potentielle. La flore y est généralement une version appauvrie de la chênaie calcifuge, en raison d’une feuillaison précoce qui limite la lumière parvenant au sol et d’une litière à décomposition lente ;
– les bois de Robinier : le Robinier Robinia pseudacacia est un arbre d’origine nord-américaine introduit en France en 1601 à des fins ornementales et aujourd’hui largement naturalisé, le plus souvent dans des sites rudéralisés (talus et délaissés routiers, voies ferrées, lisières anthropisées et coupes de massif forestiers). Essence pionnière s’accommodant de sols très variés, le Robinier possède, comme l’Aulne glutineux, des racines et des nodosités capables de fixer l’azote atmosphérique. Son bois est un excellent combustible et possède de très bonnes propriétés mécaniques. Les robiniaies « spontanées » forment généralement des bosquets linéaires et/ou de faible étendue, alors que les robiniaies plantées (4500 ha en Poitou-Charentes en 2009) occupent de petites parcelles disséminées au sein des massifs ; les peuplements y sont clairs et la strate herbacée, très fournie, comprend de nombreuses nitrophytes. La variabilité de la flore est importante et dépend en partie de la réaction du sol (robiniaies sur sols acides/r.sur sols alcalins) ;
– les bois de Charme : le Charme Carpinus betulus est une essence indigène en Poitou-Charentes (plus commune toutefois dans la moitié est de la région) qui participe à divers habitats forestiers relevant du FRAXINO-QUERCION, en compagnie des chênes, des frênes, des érables, des tilleuls, du merisier, sur des sols profonds et riches, calcaires à modérément acides. Son bois, dense et très dur, le rend apte à diverses utilisations spécialisées (tournerie, pièces de machine) et produit un combustible de qualité. Ces diverses qualités, associées au fait que l’arbre rejette fortement de souche après coupe et tolère bien la taille, font qu’il est parfois planté en peuplements purs sur de petites surfaces. Ces charmilles produisent des peuplements assez sombres et généralement homogènes quant aux classes d’âge, dont la flore n’est qu’une variante appauvrie de la chênaie-charmaie régionale ;
– les bois de Tremble : le Tremble Populus tremula est une essence pionnière à enracinement superficiel, de faible longévité (moins d’un siècle), rejetant de souche et drageonnant abondamment. Il prospère sur des sols variés – argiles, limons, sables -, basiques ou acides, mais souvent plus ou moins hydromorphes (sols mouilleux). Très rarement planté en raison de la médiocrité de son bois comme combustible et d’anciens usages pour la pâte à papier ou les allumettes aujourd’hui assumés par les peupliers euraméricains, beaucoup plus productifs, il forme parfois de petits bosquets dans les forêts dégradées (après coupes ou incendies) dont il constitue une étape de la cicatrisation. Son houppier clair laissant passer la lumière permet à une flore de sous-bois assez diversifiée de se développer (variable en fonction du degré d’engorgement du sol, de la densité des drageons etc.) ;
– les bois de Bouleau : le Bouleau verruqueux Betula pendula est une essence pionnière, héliophile, à croissance rapide et faible longévité. Très rustique, il s’accommode d’une grande variété de sols, avec une prédilection toutefois pour les substrats acides. Il fournit un bon combustible et son bois se prête bien à la fabrication de pâte à papier ou de contre-plaqué alors que son écorce d’un blanc argenté et son feuillage virant au jaune d’or en automne lui confèrent d’indéniables qualités ornementales. Les bois de bouleau se présentent le plus souvent sous la forme de petits bosquets clairs participant à la reconstitution d’une forêt acidiphile après coupe ou incendie, ou à la dynamique naturelle de vieillissement d’une lande à bruyères. Plus rarement, ils s’observent en situation pionnière dans des sites rudéralisés (carrières abandonnées, y compris sur calcaire !, friches urbaines ou industrielles). La faiblesse du couvert favorise le maintien d’une strate herbacée peu différente des habitats voisins, mêlant des éléments de landes, de fourrés calcifuges hauts à Ajonc ou Genêt où finissent par s’implanter tôt ou tard les semences des essences post-pionnières qui vont constituer la future forêt (glands enterrés par les micromammifères, ou divers Corvidés) ;
– les bois d’Orme : l’Orme champêtre Ulmus campestris s.lato appartient à un groupe de petites espèces dont la systématique est peu claire et dont le statut régional est encore largement inconnu. Dans tous les cas, il s’agit d’une essence nomade, à longévité élevée (jusqu’à 500 ans), affectionnant les sols riches en bases et en azote, à bonne alimentation en eau. Son bois, très apprécié autrefois comme combustible, est aujourd’hui réservé à des usages nobles comme l’ébénisterie ou la tournerie. Très largement planté autrefois autour des habitations – et parfois conduit en têtard par l’émondage régulier (le feuillage était donné en pâture au bétail) – l’orme forme encore de nombreux bosquets dans le paysage rural régional. Ces ormaies, souvent défigurées par la graphiose – une maladie cryptogamique causée par un champignon microscopique qui se propage dans les galeries d’un insecte le Scolyte de l’orme Scolytus scolytus – présentent une structure très variable selon leur localisation et leur état sanitaire ; seules ou associées à d’autres essences nomades et plastiques comme les frênes ou les érables, elles forment le plus souvent des haies linéaires qui, dans certains secteurs, contribuent à la formation d’un bocage plus ou moins structuré. Leur flore associée peut, selon les cas, être très nitrophile (ormaies rudéralisées, péri-urbaines) ou, au contraire, inclure diverses mésophytes forestières comme le Lierre, l’Iris fétide, l’Arum d’Italie ou le Fragon.
Phytosociologie et correspondances typologiques
PVF 2004
Les 6 sous-types s’insèrent dans la grande classe des QUERCO ROBORIS-FAGETEA SYLVATICAE Br.-Bl. & Viegler 1937 (forêts tempérées caducifoliées). Certains, comme les châtaigneraies et les bétulaies, sont plus liés aux communautés acidophiles des Quercetalia roboris Tüxen 1931, les autres aux forêts acidiclines à calcicoles des Fagetalia sylvaticae Pawlovski 1928.
Les robiniaies, les tremblaies et les ormaies peuvent aussi dans certains cas participer à des manteaux ou fruticées des CRATAEGO MONOGYNAE-PRUNETEA SPINOSAE Tüxen 1962.
COR 1991
- 41.A Bois de charmes
- 41.B bois de bouleaux
- 41.D Bois de Tremble
- 41.F Bois d’ormes
- 41.H Autres bois caducifoliés
Directive Habitats 1992 et Cahiers d’habitats
Nc.
Confusions possibles
Chacun des sous-types étant défini par la dominance physionomique d’une essence spécifique particulière, leur reconnaissance ne pose pas de problème particulier. Comme toujours en milieu naturel, on peut toutefois être confronté à des situations de transition – taillis de châtaigniers sous une futaie plus ou moins dense de chênes, accrus de bouleaux au sein d’une chênaie acidophile, faciès à orme en lisière d’une chênaie-frênaie etc. Dans tous ces cas, l’attribution à l’un ou l’autre des habitats est affaire d’échelle de perception : si l’on se réfère à la dynamique forestière, on peut considérer n’être en présence que d’un sylvofaciès de la forêt potentielle (= faciès à Betula pendula d’une chênaie acidophile) ; si on veut au contraire souligner l’originalité structurale et spécifique du faciès (car après tout un bois de bouleaux n’a pas la même structure et n’abrite pas les mêmes cortèges d’espèces qu’une chênaie), rien n’empêche de l’isoler d’une manière ou d’une autre (cartographie, dénomination phytosociologique ..).
Dynamique
Ces boisements s’insèrent dans la dynamique de la série forestière à laquelle ils participent, très variable selon les sous-types et les situations (cf. éléments dans le & « Physionomie-Ecologie »).
Espèces indicatrices
| [plante2] | Betula pendula, Carpinus betulus, Castanea sativa, Populus tremula, Robinia pseudacacia, Ulmus campestris |
| [champignons] | Châtaigniers : Russula violeipes fo. citrina
Bouleaux : Lactarius glycyosmus, Lactarius necator, Lactarius pubescens, Lactarius torulosus, Leccinum molle, Leccinum scabrum, Leccinum versipelle, Paxillus involutus, Russula betularum, Russula versicolor |
Valeur biologique
Aucun des habitats décrits ci-dessus ne présente un quelconque caractère de rareté. Pour certains (bétulaie, tremblaie), leur intérêt réside dans le fait qu’ils représentent une étape obligatoire de la reconstitution d’une forêt mature après sa dégradation ou un évènement catastrophique (incendie). Pour les autres, leur intérêt d’un point de vue de la biodiversité dépend étroitement de leur structure (boisements linéaires ou spatiaux), de leur histoire (sites fortement anthropisés/sites naturels), de leurs contacts (boisements isolés/boisements jouxtant des massifs forestiers) et, bien sûr du type de gestion sylvicole pratiquée (taillis à courte révolution/maturation avec conservation de bois morts et d’arbres sénescents).
La flore associée à ces boisements n’abrite en principe pas d’espèce rare ou menacée.
Ils n’hébergent pas d’éléments spécifiques de faune vertébrée : les mammifères et l’avifaune y sont semblables à celles des séries forestières auxquelles ils se rattachent. En ce qui concerne les invertébrés, le Robinier et le Châtaignier n’étant pas des essences indigènes, leurs communautés associées sont beaucoup plus pauvres que celles du Tremble, du Charme ou de l’Orme.
Menaces
Aucune menace particulière ne pèse sur ces habitats. On signalera seulement que le Châtaignier et l’Orme sont sensibles à diverses maladies cryptogamiques (encre et chancre pour le premier, graphiose pour le second) qui peuvent compromettre la qualité et la quantité des produits récoltés mais ne semblent par particulièrement nuisibles à la biodiversité globale de ces habitats.
Statut régional
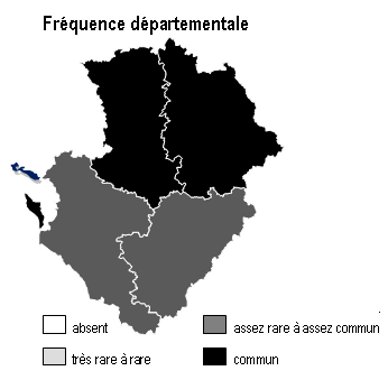
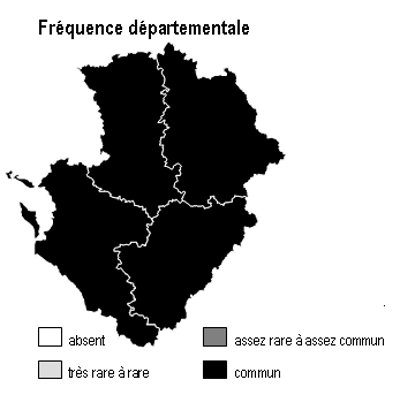
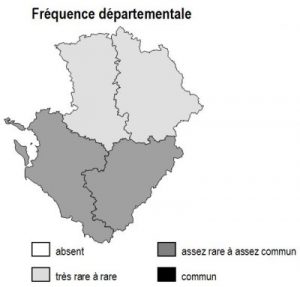
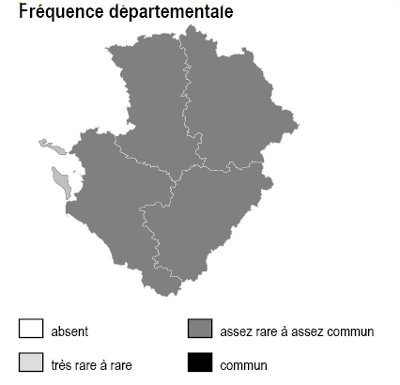
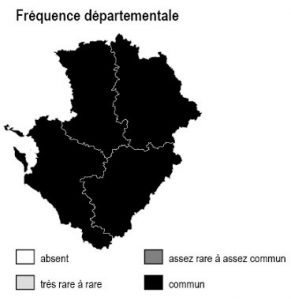
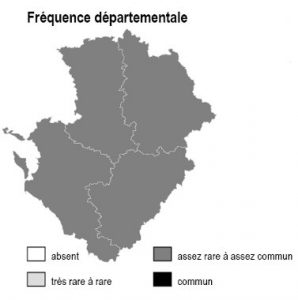
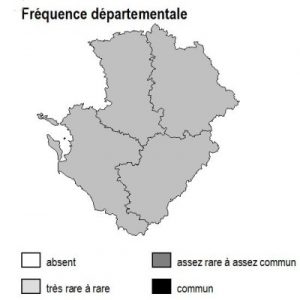
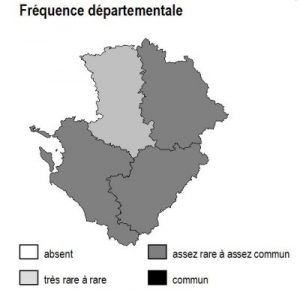
Les 6 sous-types existent dans chacun des 4 départements mais avec des abondances variables selon la nature des sols :
– bois de châtaigniers : ils couvrent des surfaces importantes sur les argiles à silex et les terres rouges du nord 16, sud-est 79, sud-ouest 86 ainsi qu’en Saintonge centrale (17) et dans les 2 Doubles (16 et 17)
– bois de robiniers : ils sont surtout répandus en 86, notamment sur les sables du Loudunais, du Saumurois, ainsi qu’en plaine de Thouars (79)
– bois de charmes : ils sont fréquents dans une grande partie de 86, sur les terres rouges de 79, la marge orientale de 16 et en Saintonge centrale
– bois de trembles : ils sont dispersés partout sans pattern particulier
– bois de bouleaux : ils sont fréquents sur les terrains non calcaires : Basse-Marche de 86 et 16, bocage et hauteurs de Gâtine en 79, ainsi qu’à un degré moindre dans la Double (16 et 17)
– bois d’orme : ils sont présents à peu près partout